
- Prinsip Dasar: Perpasangan Basa ke Untai Cetakan
- Sintesis Untai DNA Baru
- Replikasi Ujung-Ujung Molekul DNA
Prinsip Dasar: Perpasangan Basa ke Untai Cetakan
Watson dana Crick menyatakan hipotesis tentang cara DNA bereplikasi:
Kini model kami tentang asam deoksiribonukleat merupakan, sebagai akibatnya, sepasang cetakan, masing- masing saling komplementer. Kami membayangkan bahwa sebelum duplikasi, ikatan hidrogen putus, dan kedua rantai membuka serta memisah. Setiap rantai kemudian bertindak sebagai cetakan untuk pembentukan rantai pendamping pada dirinya sendiri, sehingga kita akhirnya akan memperoleh dua pasang rantai, yang awalnya hanya satu. Terlebih lagi, urut-urutan pasangan basa akan diduplikasi secara tepat sama.*
Peraga 16.9 mengilustrasikan gagasan dasar Watson dan Crick. Agar lebih mudah diikuti, kami hanya menunjukkan bagian pendek heliks ganda dalam bentuk yang tidak terulir. Perhatikan bahwa jika Anda menutupi salah satu untai DNA pada Peraga 16.9a, Anda masih dapat menentukan urutan linear basa-basa di untai tersebut dengan mengacu kepada untai yang tidak ditutupi dan menerapkan aturan perpasangan basa. Kedua untai tersebut bersifat komplementer. Setiap untai menyimpan informasi yang diperlukan untuk merekonstruksi untai yang satu lagi. Ketika setiap sel menyalin suatu molekul DNA, setiap untai berperan sebagai cetakan untuk mengurutkan nukleotida menjadi untai baru yang komplementer. Nukleotida berjejer di sepanjang untai cetakan berdasarkan aturan perpasangan basa dan ditautkan membentuk untai baru.

Pada awal proses hanya ada satu molekul DNA yang beruntai ganda, namun kemudian ada dua, masing-masing merupakan replika yang sama persis dengan molekul ‘induk. Mekanisme penyalinan ini analog dengan negatif foto untuk membuat citra positif, yang kemudian juga bisa digunakan untuk membuat negatif foto yang lain, dan demikian seterusnya.
Model replikasi DNA ini tetap tidak teruji selama beberapa tahun setelah publikasi struktur DNA. Percobaan yang diperlukan bersifat sederhana secara konsep, namun sulit dilaksanakan. Model Watson dan Crick memprediksikan bahwa saat heliks ganda bereplikasi, masing-masing dari kedua molekul anakan mengandung satu untai lama, yang berasal dari molekul induk, dan satu untai yang baru dibuat. Model semikonservatif (semiconservative model) ini dapat dibedakan dari model replikasi konservatif, yang menyatakan kedua untai induk kembali menyatu setelah proses tersebut (artinya, molekul induk akan dilestarikan). Dalam model yang ketiga, disebut model dispersif, keempat untai DNA yang ada setelah replikasi merupakan campuran DNA lama dan baru (Peraga 16.10).

Sintesis Untai DNA Baru
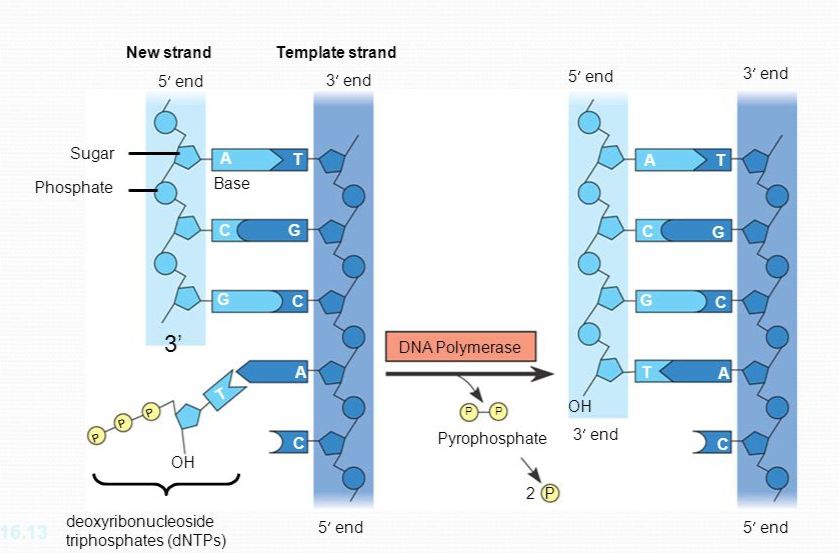
Enzim-enzim yang disebut DNA polimerase (DNA polymerase) mengkatalisis sintesis DNA baru dengan cara menambahkan nukleotida-nukleotida ke rantai yang telah ada sebelumnya. Pada E. coli, ada beberapa DNA polimerase yang berbeda, namun tampaknya ada dua yang berperan utama dalam replikasi DNA: DNA polimerase III dan DNA polimerase I. Situasi pada eukariota lebih rumit dengan setidaknya 11 DNA polimerase yang telah ditemukan sejauh ini. Akan tetapi, prinsip-prinsip umumnya tetap sama.
Sebagian besar DNA polimerase membutuhkan primer dan untai cetakan DNA, selain jejeran nukleotida DNA komplementer. Pada E. coli, DNA polimerase III (disingkat DNA pol III) menambahkan suatu nukleotida DNA ke primer DNA dan kemudian meneruskan penambahan nukleotida DNA, yang komplementer dengan untai cetakan DNA induk, ke ujung untai baru DNA yang sedang tumbuh. Laju pemanjangan kira-kira 500 nukleotida per detik pada bakteri dan 50 per detik pada sel manusia.
Setiap nukleotida yang ditambahkan ke untai DNA yang sedang tumbuh berasal dari nukleosida trifosfat, yang merupakan suatu nukleosida (gula dan basa) dengan tiga gugus fosfat. Anda sudah pernah menjumpai molekul semacam itu-ATP (adenosin trifosfat, lihat Peraga 8.8). Satu-satunya perbedaan antara ATP metabolisme energi dan dATP, nukleosida trifosfat yang memberikan nukleotida adenin ke DNA, adalah komponen gulanya. Gula yang digunakan pada balok pembangun DNA berupa deoksiribosa, sedangkan pada ATP berupa ribosa. Seperti ATP, nukleosida trifosfat yang digunakan untuk sintesis DNA reaktif secara kimiawi, sebagian karena ekor trifosfat memiliki gugus muatan negatif yang tak stabil. Ketika setiap monomer bergabung ke ujung untai DNA yang sedang tumbuh, dua gugus fosfat hilang sebagai suatu teolekul pirofosfat – Hidrolisis pirofosfat yang terjadi setelahnya menjadi dua molekul fosfat anorganik i adalah reaksi eksergonik tergandengkan yang membantu menggerakkan reaksi polimerisasi (Peraga 16.14).
Replikasi Ujung-Ujung Molekul DNA
Ada sebagian kecil DNA sel yang tidak dapat direplikasi atau diperbaiki oleh DNA polimerase. Untuk DNA lurus, misalnya DNA kromosom eukariot, fakta bahwa DNA polimerase hanya dapat menambahkan nukleotida ke ujung 3′ polinukleotida yang sudah ada sebelumnya menyebabkan masalah. Mekanisme replikasi yang biasa tidak memberikan cara untuk menyelesaikan ujung-ujung 5′ untai DNA anakan. Meskipun fragmen Okazaki bisa dimulai dengan primer RNA yang berikatan dengan bagian paling ujung dari untai cetakan, begitu dibuang, primer tidak dapat digantikan oleh DNA karena tidak ada ujung 3′ yang tersedia untuk penambahan nukleotida (Peraga 16.19). Akibatnya, putaran demi putaran replikasi yang berulang-ulang menghasilkan molekul-molekul DNA yang semakin pendek dengan ujung tak rata.
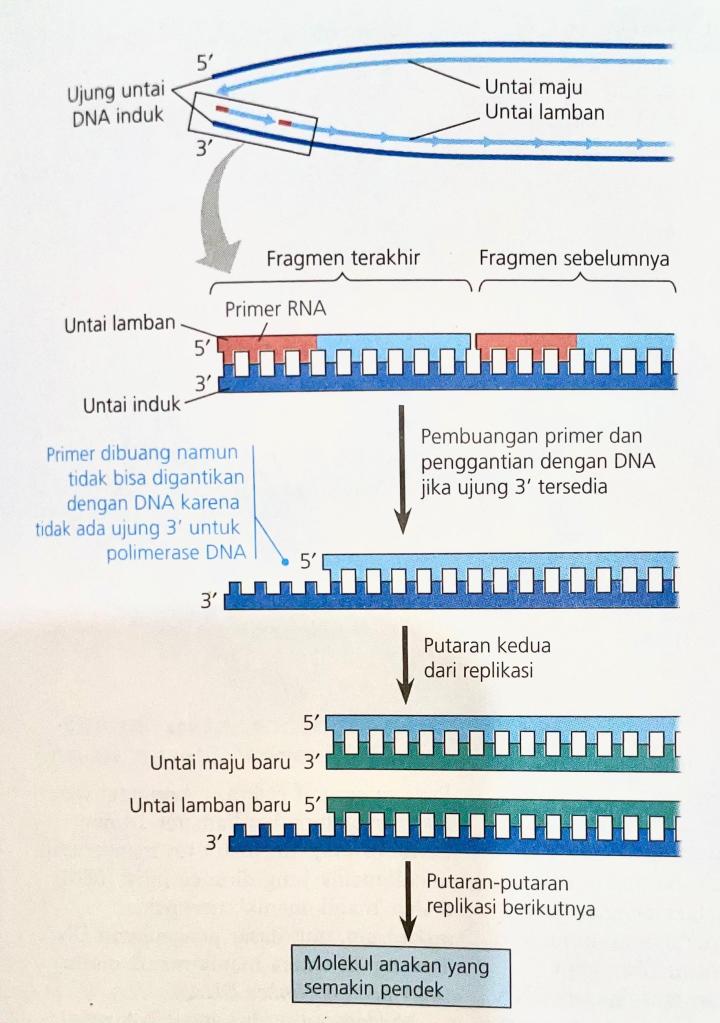
Pemendekan DNA tidak terjadi pada sebagian besar prokariota karena DNA prokariot berbentuk melingkar, sehingga tidak memiliki ujung. Namun, apa yang mencegah gen-gen eukariot terkikis habis meskipun replikasi DNA terjadi berulang-ulang.Ternyata molekul DNA kromosom eukariot memiliki sekuens nukleotida khusus yang disebut telomer (telomere) di bagian ujung (Peraga 16.20). Telomer tidak mengandung gen. Sebagai gantinya, DNA umumnya mengandung banyak pengulangan satu sekuens nukleotida pendek. Pada setiap telomer manusia, misalnya, sekuens enam nukleotida TTAGGG diulang- ulang sebanyak 100 sampai 1.000 kali. DNA telomer melindungi gen-gen organisme. Selain itu, protein spesifik yang terkait dengan DNA telomer mencegah ujung somplak dari molekul anakan mengaktivasi sistem-sistem sel untuk mengawasi kerusakan DNA. (Ujung somplak molekul DNA, yang sering disebabkan oleh patahan untai ganda, dapat memicu jalur-jalur transduksi sinyal yang menyebabkan penghentian siklus sel atau kematian sel. Sejenis enzim bernama Telomerase mengaktualisasikan pemanjangan telomer dalam sel-sel nutfah eukariot, sehingga mengembalikan panjang awal dan mengompensasi pemendekan yang terjadi selama replikasi DNA. Telomerase tidak aktif pada sebagian besar sel somatik manusia, namun aktivitasnya dalam sel nutfah menghasilkan telomer dengan panjang maksimum pada zigot.
Pemendekan normal telomer mungkin melindungi organisme dari kanker dengan cara membatasi jumlah pembelahan yang bisa dialami oleh sel somatik. Sel-sel dari tumor besar seringkali memiliki telomer yang pendeknya tak biasa, seperti yang bisa diduga dari sel-sel yang telah mengalami banyak pembelahan sel. Pemendekan lebih lanjut mungkin menyebabkan penghancuran diri sel tumor. Para peneliti ternyata telah menemukan aktivitas telomerase dalam sel-sel somatik yang menjadi kanker. Hal tersebut menunjukkan bahwa kemampuan aktivitas enzim tersebut tampaknya menstabilkan panjang telomer sehingga sel-sel kanker bisa terus membelah. Banyak sel kanker tampaknya mampu membelah tanpa terbatas, seperti pada galur imortal sel-sel kultur (lihat Bab 12). Jika telomerase ternyata memang faktor penting pada banyak kanker, mungkin telomerase dapat menjadi target yang berguna untuk diagnosis maupun kemoterapi kanker.

